

圖│研之有物
找到原創性主題
上篇〈看真菌如何獵殺線蟲,開啟寄生蟲治療藥物新曙光!〉文章介紹,薛雁冰團隊發現微觀世界的狩獵現場:線蟲捕捉菌設陷阱、杏鮑菇下毒捕食線蟲,這些殺蟲機制將對生物防治帶來全新的可能。但更寶貴的是,在薛雁冰開始研究這個課題時,科學家對於食蟲真菌和線蟲互動的這些分子機制,所知甚少,她可說是開路先鋒。
「科學家估計地球上的真菌約有五百萬種,但目前研究者真正了解的,只有酵母菌、麵包黴及一些病原菌等十多種真菌。但自然界還有一些非常有趣的真菌,幾乎沒有分子層次的研究。」薛雁冰感性地回憶:「非常感謝我在加州理工學院博士後研究的指導教授 Paul Sternberg 願意支持我的想法,讓我在他的實驗室中嘗試開啟一個新領域,利用模式線蟲C. elegans 來探討食蟲真菌跟線蟲之間的獵食者-獵物的交互作用,分子機制和共同演化。」回到台灣,在中研院建立自己的實驗室之後,薛雁冰繼續帶領著團隊,拓展食蟲真菌的研究,一步步拼湊出食蟲真菌獵殺線蟲的分子機制。
但要當個開路先鋒可不容易,從找到有價值的原創性主題、建立更好的模式生物,甚至打造分析工具,每往前一步皆須面臨更大的未知。但也因原創性的貢獻,研究成果頻頻登上如《美國國家科學院院刊》 (PNAS) 等國際期刊。一起走進薛雁冰團隊的研究現場!
野外採集,驗證實驗非空想
薛雁冰團隊發現線蟲捕捉菌 A.oligospora 會透過五個步驟捕食 C.elegans 線蟲:吸引獵物 → 發現獵物 → 設下陷阱 → 抓住獵物 → 飽餐一頓。
BUT!在大自然中,線蟲和線蟲捕捉菌真的會碰在一起,並如在實驗室般「打得火熱」嗎?於是薛雁冰團隊穿梭在臺灣山野間進行採集,結果發現,臺灣三分之二的土壤均有線蟲捕捉菌,而線蟲又是數量最多的動物,兩者果真常常「同在一起」。

圖│研之有物 (資料來源│薛雁冰)
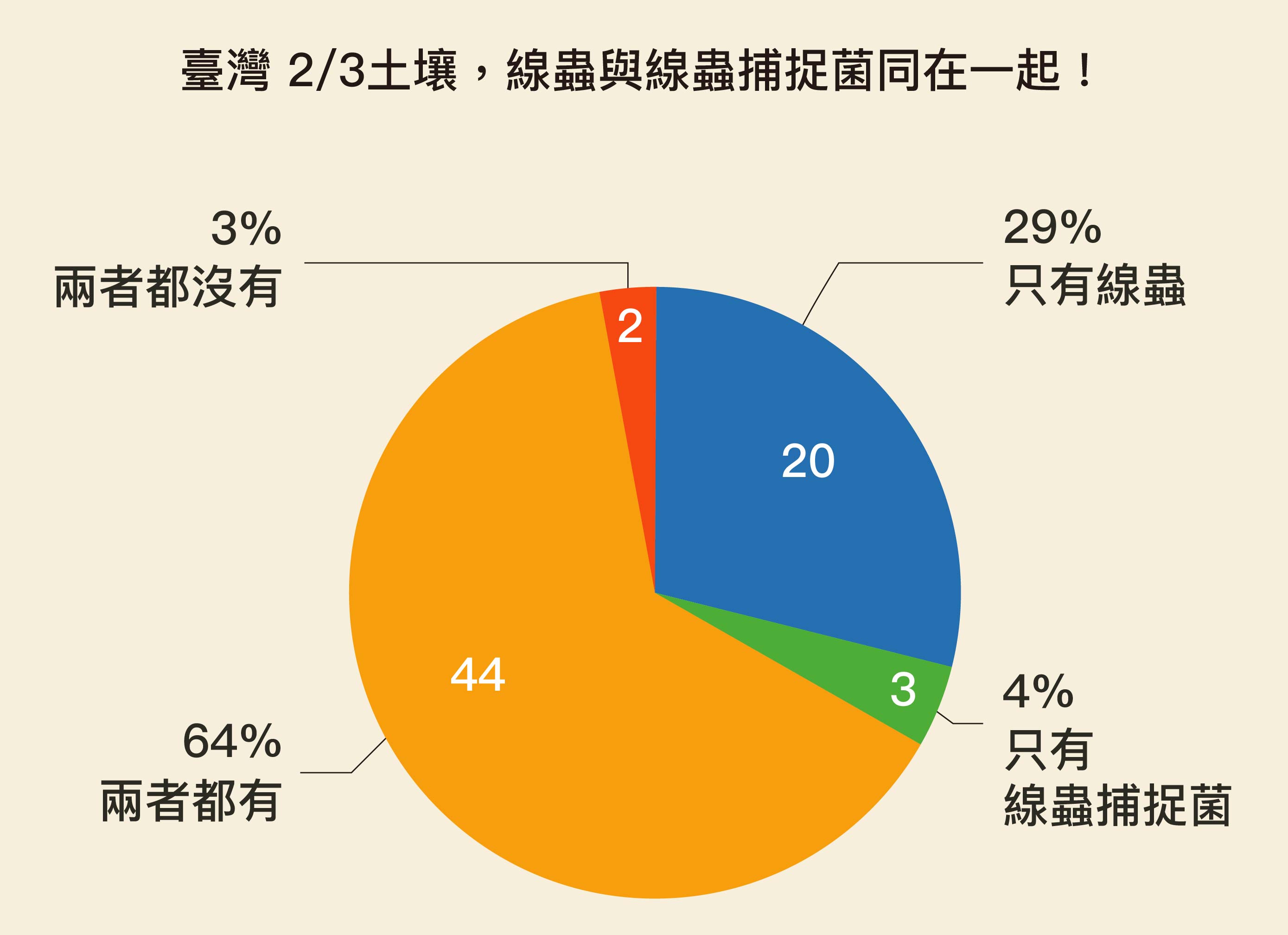
他們也從泥土中分離出各種線蟲和真菌,整理在大自然中哪些線蟲和哪些線蟲捕捉菌生長在一起,在實驗室測試牠們的互動,比方說:某種線蟲捕捉菌是不是什麼線蟲都吃?還是只吃幾種線蟲?結果發現,線蟲捕捉菌「不挑吃」,什麼線蟲都吃,狩獵關係果真「普遍存在」所有的線蟲與線蟲捕捉菌之間。
發現超強菌種,建立研究好素材
研究過程中,薛雁冰也發現從菌種中心獲得的線蟲捕捉菌標準菌株「有點弱弱的」,不利於研究。怎麼辦?自己建立新模型!他們從野外採集的線蟲捕捉菌中,找到一些狩獵能力較強菌株。這些菌株可以產生很多捕捉構造,而且長出捕捉構造的速度比較快,因此殺死線蟲的速度也快!這些超強菌株可以成為絕佳的模式菌株,讓科學家們更容易研究線蟲捕捉菌如何長出捕捉構造,捕捉獵物需要哪些基因。
薛雁冰也以基因體定序加上基因體剔除法,破解超強菌株的狩獵力秘密:G 蛋白!在線蟲捕捉菌身上,G 蛋白負責對細胞傳遞外界的訊號,薛雁冰發現:這個蛋白會去傳遞外界有線蟲出沒的訊號,促使細胞長出捕捉構造。如果沒有這個蛋白,線蟲捕捉菌就無法發育出捕捉構造,只能讓「到嘴的肥蟲」逃走囉~
戰鬥防禦力分析工具
線蟲和真菌的實驗樣本要耐心尋找,分析它們戰鬥防禦力的工具也要費心開發。如果你曾經把麵包放到發霉,有天驀然回首,你會發現麵包長出一塊一塊的黴菌菌絲,很難精準描述其生長速度、生長範圍、菌絲長度。實驗室裡的線蟲捕捉菌(真菌)也是類似的情況。
在薛雁冰實驗室中,博士後研究員 Guillermo Vidal-Diez 透過電腦影像視覺分析的技術,自行發展出可以「定量」描述真菌生長的工具,如下圖所示:
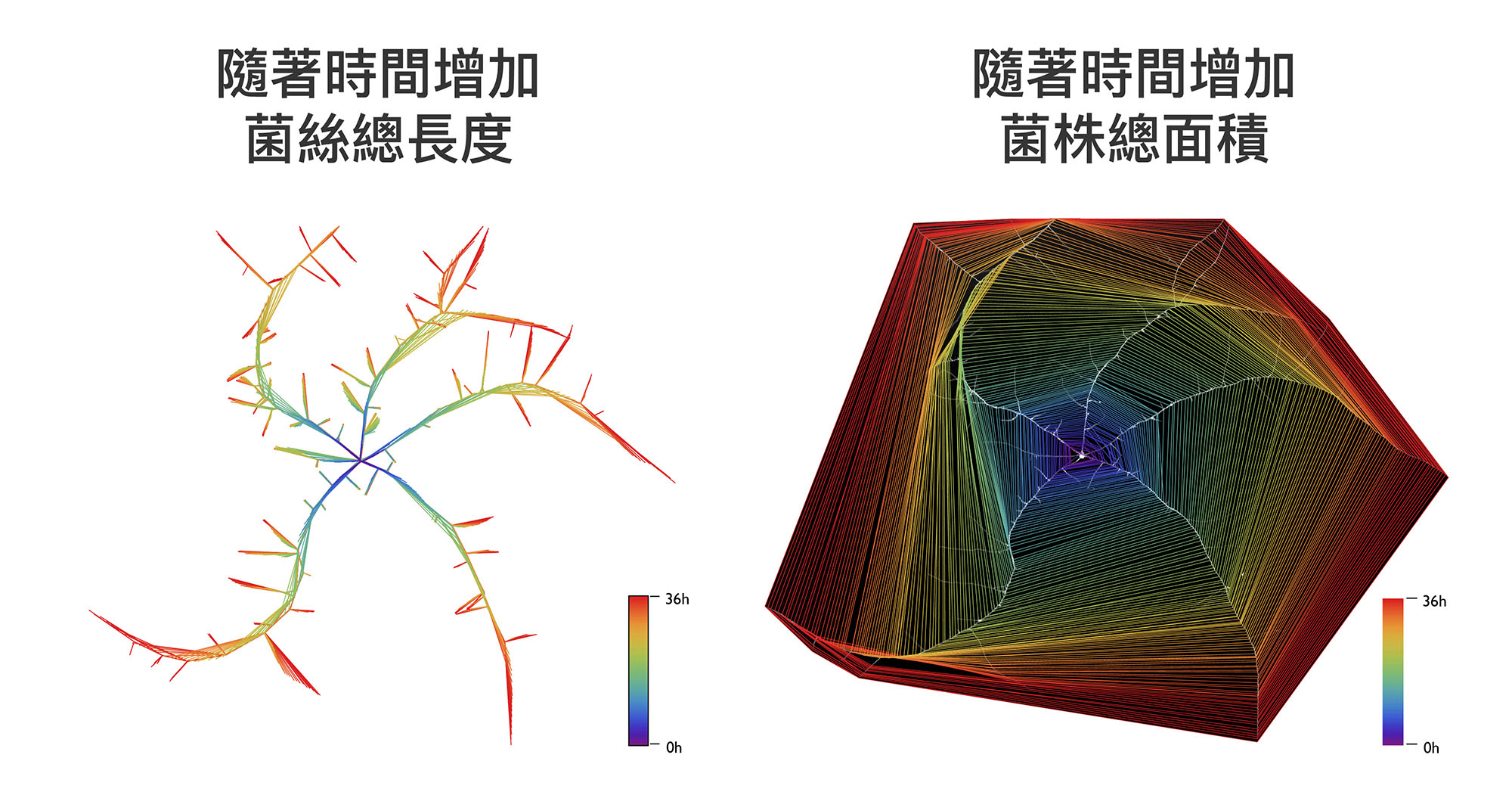
圖│Guillermo Vidal-Diez de Ulzurrun,Tsung-Yu Huang,Ching-Wen Chang,Hung-Che Lin,Yen-Ping Hsueh(2019) Fungal feature tracker (FFT): A tool for quantitatively characterizing the morphology and growth of filamentous fungi. PLoS Comput Biol. 15 (10) : e1007428
隨著科學技術的快速發展,現在做研究比以前快上許多。例如,以前要花上數年才能找出突變線蟲的基因變異所在,現在只要一兩個月,就能藉由遺傳學的分析、加上全基因體定序,快速找出是哪些基因發生變異,造成這個突變線蟲的性狀改變 (例如外型、神經系統的發育,或是行為發生改變),因此能躲過線蟲捕捉菌的吸引和陷阱,逃過被捕食的命運。
「這些快速發展的技術和工具,提供我們一個很好的時代,再次利用『遺傳學的強大力量』(the awesome power of genetics) 來研究生物學上重要的問題,」薛雁冰說:「這些工具讓我有勇氣去探索『非模式物種』 (non-model organisms) 的奧秘,開拓新領域來建立起我們的原創性研究課題。」





